
Figure 5 : Voies nerveuses du réflexe photomoteur. D’après Adams et Victor [1].
Physiologie pupillaire
Physiologie de la motilité pupillaire
La motilité irienne est influencée par trois ordres de facteurs qui s’intriquent constamment :
- des facteurs en rapport avec la fonction visuelle qui régissent le réflexe pupillo-moteur direct et consensuel, la réaction pupillaire à la vision rapprochée, et, plus accessoirement, la réaction syncinétique de Piltz et Westphal de la contraction de l’orbiculaire ;
- des facteurs en rapport avec l’activité des centres cérébro-spinaux expliquant : les réflexes sensitivo-moteurs, les réflexes sensori-moteurs, les réflexes psycho-moteurs ;
- des facteurs humoraux qui sont cholinergiques pour toutes les fibres pré-ganglionnaires qu’elles soient sympathiques ou parasympathiques. C’est aussi l’acétylcholine qui est libérée au niveau des fibres post-ganglionnaires du parasympathique tandis que l’adrénaline intervient au niveau des fibres post-ganglionnaires du système sympathique. Le point d’impact des différentes substances susceptibles d’influencer le diamètre pupillaire est variable, expliquant les modalités précises d’action des différentes substances parasympathomimétiques, parasympatholytiques, sympathomimétiques et sympatholytiques [29].
Réflexes
pupillaires normaux
Echappant au contrôle volontaire, la pupille est régie sur un mode réflexe.
La contraction pupillaire sous l’effet de la lumière est normalement rapide, se maintient le temps de l’excitation lumineuse, elle est suivie d’une décontraction, également rapide, dès que l’illumination cesse : c’est le réflexe photomoteur direct. L’éclairement d’une rétine entraîne la contraction pupillaire controlatérale : c’est le réflexe consensuel chaque rétine ayant des connexions avec les noyaux oculomoteurs des deux côtés [28,39]. La latence du réflexe photomoteur normal est comprise entre 200 et 250 ms [28].
La voie du réflexe photomoteur, qui aboutit à une irido-constriction comporte quatre neurones [52]. Les photorécepteurs rétiniens transmettent leurs informations au premier neurone de la voie afférente, dit neurone pupillaire, dont l’axone suit la voie optique. Dans le chiasma optique, les fibres pupillaires se divisent en deux contingents, droit et gauche (réflexe photomoteur consensuel) ;elles parcourent ensuite les bandelettes optiques. Puis, sans avoir atteint le corps genouillé externe, elles empruntent le bras conjonctival du tubercule quadrijumeau antérieur. Elles font alors relais dans la région prétectale avec le deuxième neurone, au niveau des deux noyaux du tractus optique et des deux noyaux olivaires [1,39,52].
Les fibres pupillaires gagnent ensuite, selon un court trajet, le noyau d’Edinger-Westphal homolatéral, ainsi que le noyau controlatéral par l’intermédiaire de fibres qui traversent la commissure postérieure [1,39,52].
D’autres connexions entre le cortex et les centres pupillo-moteurs existent en outre, dessinant la voie supra-nucléaire du réflexe : l’aire 19 en est le relais cortical, dont l’efférence se projette sur le noyau d’Edinger-Westphal. Cette voie contrôle la syncinésie accommodation-convergence [28].
Enfin, à partir de ce noyau, la voie parasympathique oculaire à deux neurones constitue la branche efférente motrice du réflexe [1,39,52].
Le schéma suivant (figure 5) retrace le trajet de cette voie réflexe.
Figure 5 : Voies nerveuses du réflexe photomoteur. D’après Adams et Victor [1].
Le réflexe de mydriase consécutif à l’obscuration ne semble pas être un simple phénomène passif, par relâchement du sphincter irien. Il associe probablement, à partir des récepteurs rétiniens, une inhibition de la voie du réflexe photomoteur dans le mésencéphale et une stimulation de la voie sympathique de l’irido-dilatation, selon un arc réflexe encore mal connu. En effet, après section du sympathique, la mydriase consécutive à l’obscuration est moins prononcée. Les voies anatomiques de la dilatation pupillaire après stimulation sensitive, sensorielle ou psychique sont encore moins connues [52].
La constriction pupillaire qui accompagne l’accommodation-convergence (ou réaction pupillaire à la vision de près) [28,39] n’est pas un vrai réflexe, mais un mouvement syncinétique d’origine supra-nucléaire. C’est donc une synergie de trois mécanismes dite “triade de fixation” : l’accommodation qui permet la mise au point sur la rétine, la convergence des globes oculaires qui assure la fusion des images rétiniennes, le myosis qui réduit les aberrations dues aux changements de courbure cristallinienne. La constriction du sphincter irien et du muscle ciliaire d’une part, et celle des muscles droits internes d’autre part interviennent alors dans cette action synergique [52].
La contraction pupillaire débute lorsque l’objet fixé est à 40 cm et devient maximale à 30 cm. Elle ne dépend ni de l’accommodation, ni de la convergence, mais de la vision de près [28,39].
Les voies de la réaction d’accommodation partent de la rétine, suivent les voies optiques jusqu’au cortex visuel, qui se projette sur les noyaux antéro-médians. Elles suivent ensuite la voie finale commune parasympathique pour gagner le ganglion ciliaire [52]. Le détail du circuit anatomique cortical et de son fonctionnement n’est pas encore connu [39].
Il existe d’autres réflexes pupillaires [28,39] :
- le réflexe sensitivo-moteur : toute excitation d’un nerf sensitif (sauf le nerf trijumeau) entraîne une mydriase ;
- le réflexe sensori-moteur : toute sensation (gustative, tactile, auditive) désagréable entraîne une mydriase bilatérale ;
- le réflexe psychomoteur : les excitations psychiques (peur, émotion, joie) provoquent une mydriase bilatérale ;
- le réflexe à l’attention de Haab (myosis léger ipsilatéral) ;
- la réaction pupillaire syncinétique à l’occlusion des paupières de Piltz-Westphal (myosis ipsilatéral) ;
- le réflexe pupillo-constricteur à point de départ trigéminé, observé lors de toute irritation du segment antérieur du globe oculaire.
Les voies anatomiques de la dilatation pupillaire après stimulation sensitive, sensorielle ou psychique ne sont pas encore précises [52].
Remarque
Chez l’homme la mydriase provoquée par une émotion a pour mécanisme une inhibition du centre constricteur pupillaire, puisqu’elle se produit encore sur une pupille sympathectomisée alors qu’elle disparaît après section du nerf moteur oculaire commun [39].
Médiateurs
chimiques de la motilité pupillaire. Epreuves pharmacodynamiques. Epreuve des collyres.
Que ce soit dans le système parasympathique ou dans le système sympathique, la transmission du signal nerveux se fait par l’intermédiaire d’un médiateur chimique au niveau du relais ganglionnaire et à la jonction entre le neurone post-ganglionnaire et l’organe effecteur. Deux médiateurs ont été individualisés : l’acétylcholine au niveau de la synapse ganglionnaire des systèmes ortho- et parasympathique et à l’extrémité du deuxième neurone parasympathique ; la noradrénaline à la partie terminale du deuxième neurone sympathique [39].
Diverses substances pharmacologiques sont utilisées en ophtalmologie pour étudier l’intégrité des voies anatomiques. La connaissance de leur action sur une pupille normale permet alors de déterminer la voie atteinte par la lésion.
Les parasympathomimétiques (pilocarpine, ésérine) déterminent une constriction pupillaire [28].
- l’ésérine provoque un myosis et un spasme du muscle ciliaire. Son action demande quelques minutes et dure 24 à 48 heures.
- la pilocarpine à 1% stimule directement le muscle sphincter de l’iris [1], même lorsque le nerf moteur oculaire commun est paralysé. Le myosis qu’elle provoque se poursuit plusieurs heures. L’instillation de pilocarpine ne provoque pas de contraction si la mydriase est d’origine atropinique [4].
Les parasympatholytiques (belladone, atropine, homatropine) sont mydriatiques [28], par paralysie des terminaisons nerveuses parasympathiques [1] :
- le sulfate d’atropine à 1% provoque, 10 minutes après son instillation, une mydriase que la cocaïne peut encore majorer, et une abolition du réflexe photomoteur. Son action se poursuit plus de 24 heures.
- l’homatropine dilate elle aussi la pupille, de manière moins durable.
Les sympathomimétiques (cocaïne) sont également mydriatiques [28] :
- la cocaïne, produit une mydriase sur l’œil normal, en laissant persister les réflexes photomoteur et l’accommodation. Elle agit en inhibant la réabsorption dans les terminaisons présynaptiques de la noradrénaline, transmetteur libéré entre les neurones sympathiques post-ganglionnaires et les cellules du muscle dilatateur de l’iris [59].
Les propriétés de ces différentes substances pharmacologiques sont utilisées dans l’épreuve des collyres de Coppez.
L’épreuve des collyres consiste à instiller successivement dans chacun des yeux des collyres à action sympathicomimétique et parasympatholytique. L’examen des pupilles se fait alors en semi-obscurité, les deux yeux étant éclairés également [29,44].
On peut être amené à pratiquer cette épreuve dans deux circonstances [29,44]:
- lorsque les pupilles sont sensiblement égales pour faire apparaître une anisocorie peu apparente et latente ;
- devant une anisocorie, elle permet de déterminer le côté pathologique et le système (sympathique ou parasympathique) en cause.
Lorsque les pupilles sont sensiblement égales on utilise l’homatropine à 0,5%, parasympatholytique faible, instillée une fois dans les deux yeux. Les pupilles sont ensuite examinées toutes les cinq minutes, un des côtés se dilatera moins que l’autre en cas d’anisocorie.
Lorsqu’il y a anisocorie, on explore en premier la pupille la plus large avec la cocaïne à 2% comme sympathomimétique. Trois ou quatre instillations sont faites à 3 ou 4 minutes d’intervalle et on examine les pupilles toutes les 15 minutes pendant une heure.
Il y a trois possibilités de réponse [18,28,29] :
- la pupille ne se dilate pas ou peu. Il s’agit d’une mydriase par spasme du dilatateur (excitation des fibres dilatatrices sympathiques). Les fibres dilatatrices étant déjà excitées, la cocaïne, sympathomimétique ne peut plus avoir d’action sur elles (cas du syndrome de Pourfour du Petit) ;
- la pupille se dilate largement. Il s’agit d’une mydriase par paralysie du sphincter. La cocaïne conserve tout son pouvoir dilatateur et le spasme du dilatateur qu’elle produit s’ajoute à la paralysie préexistante du sphincter pour amener une mydriase maximale (en cas d’atteinte du nerf oculomoteur par exemple) ;
- la pupille se dilate un peu. Il s’agit de la pupille normale où l’excitation du dilatateur est contrebalancée par le tonus du sphincter.
On se porte ensuite vers la pupille la plus étroite qui peut être soit normale, soit le siège d’une excitation du sphincter ou d’une paralysie du dilatateur. La cocaïne est instillée comme précédemment [29] :
- la pupille se dilate. Il s’agit d’une pupille normale dont la dilatation est d’autant plus visible que son diamètre au départ était plus faible ;
- la pupille ne se dilate pas ou très peu. Elle est pathologique et pour aller plus loin, il faut avoir recours à un parasympatholytique, homatropine ou atropine à 0,5 ou 1 %. Sous l’effet de celui-ci :
- la pupille se dilate largement : il s’agit d’un myosis spasmodique levé par le parasympatholytique (excitation du parasympathique) ;
- la pupille se dilate faiblement : il s’agit d’une paralysie du dilatateur à laquelle est venue s’ajouter la privation du tonus du sphincter (syndrome de Claude Bernard-Horner).
L’instillation d’atropine dans une pupille en mydriase par excitation du muscle dilatateur provoque une dilatation maximale [18].
Le schéma suivant résume le protocole de l’épreuve des collyres.
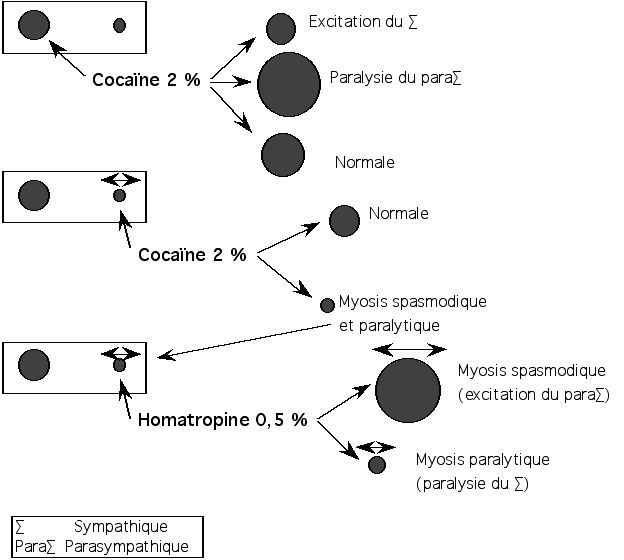
Figure 6 : Epreuves des collyres de Coppez. D’après Saraux [44]
Il faut préciser cependant que l’épreuve des collyres est parfois d’interprétation délicate et que sa technique doit être rigoureuse, en particulier dans le respect du délai (parfois plusieurs jours) entre deux collyres utilisés [47].
Données
expérimentales
Comme il a déjà été dit, le syndrome d’excitation du sympathique fut initialement observé chez l’animal par François Pourfour du Petit en 1727. Au cours de ses expériences de section du sympathique cervical dans le cou, il avait en effet remarqué lors de ses expériences sur des animaux vivants que l’excitation de ce nerf “fait devenir les yeux plus brillants, plus vifs et plus éveillés et les pupilles plus dilatées.” [26].
Claude Bernard, reprenant en partie ses travaux, constate que l’excitation du sympathique cervical provoque une dilatation pupillaire, un élargissement de la fente palpébrale, une exophtalmie, une vasoconstriction [9,12]. Dans un rapport célèbre de 1852, il décrit la totalité des signes du syndrome d’excitation du sympathique. Il obtient après section du sympathique cervical un syndrome paralytique, le syndrome de Claude Bernard-Horner, mais note que “ le galvanisme produit les effets diamétralement opposés. Si on galvanise le bout supérieur du grand sympathique divisé, tous les phénomènes qui ont dû se produire changent de face : la pupille s’élargit, l’ouverture des paupières s’agrandit. L’œil fait saillie hors de l’orbite ; d’active qu’elle était, la circulation devient faible ; la conjonctive, les narines, les oreilles, qui étaient rouges, pâlissent. On cesse la galvanisation. Tous les phénomènes primitivement produits par la section reparaissent peu à peu, pour disparaître de nouveau à une seconde application du galvanisme. On peut continuer à volonté cette expérience, la répéter autant de fois que l’on voudra, toujours ses résultats seront les mêmes […] L’élargissement qui survient par la galvanisation du bout supérieur du grand sympathique est involontaire, l’animal ne peut pas s’y opposer [...].” [9,12].
C.E. Brown-Sequard obtient des résultats comparables qu’il décrit dans un mémoire de Janvier 1854 [15] :
“Effets de l’action énergique du nerf grand sympathique, produite par l’application du galvanisme quelque temps après que ce nerf a été coupé au cou.
1° La pupille se dilate, [...]
2° Le globe oculaire, qui était tourné en dedans et enfoncé dans l’orbite, reprend sa situation normale, [...]
3° Les paupières s’ouvrent et les contractions des muscles de la narine, de la bouche et de l’oreille cessent d’exister, [...]
4° La vascularisation diminue [...] Quelquefois les vaisseaux se contractent à ce point que le calibre des petites artères et des veinules disparaît complètement [...] Quand on cesse la galvanisation, les vaisseaux ne tardent pas à se dilater de nouveau [...] Si cette application est continuée pendant un temps très long, surtout si elle est énergique, les vaisseaux, après être restés contractés cinq, six minutes ou un peu plus, commencent à se dilater lentement, et malgré la persistance de l’excitation, on les voit se dilater notablement [...]
5° La température s’abaisse en même temps que les vaisseaux se contractent pendant la galvanisation. La narine, et surtout l’oreille, après avoir repris la même température que celles du côté sain, perdent encore de un à deux degrés centigrade [...]
6° La sensibilité diminue ; cependant cette diminution est moins facile à constater que la perte de chaleur.”
C. E. Brown-Sequard complète son étude par la stimulation du sympathique cervical non sectionné. Il constate la reproduction des faits précédents mais note que quelques-uns de ces effets sont alors moins marqués [15].
L’excitation du sympathique a donc des effets strictement inverses de ceux que l’on observe après section de la chaîne cervicale. L’effet ne dure cependant que le temps de l’excitation. On observe une mydriase qui ne modifie pas les réactions pupillaires aux divers modes.
De nombreux expérimentateurs (Braunstein, Bach, Lodato) ont travaillé à la suite de Claude Bernard sur la stimulation électrique du système sympathique. Ils mettent en évidence que si l’excitation est unique, la mydriase est également passagère et que la pupille reprend aussitôt son calibre normal, après la fin de l’excitation. Lorsque l’excitation est prolongée, tantôt on observe une mydriase durable, tantôt la pupille reprend peu à peu son calibre initial, tantôt on observe des alternatives de dilatation et de rétrécissement. On observe également un élargissement de la fente palpébrale conditionné par une élévation de la paupière supérieure et un abaissement de la paupière inférieure, ainsi qu’une protrusion du globe oculaire [26,55].
En 1892 Langhy établit qu’une dilatation pupillaire peut être obtenue à partir des racines ventrales de C8, D1 et D2. Une stimulation de la moëlle cervicale au niveau de la colonne intermédio-laterale provoque une mydriase et une réaction vasomotrice [58].
Chez l’homme, au cours d’interventions mettant à découvert le sympathique cervical, on a pu obtenir par excitation électrique une dilatation pupillaire isolée, commençant quelques secondes après le début de la stimulation, se faisant par à-coups et persistant encore un certain temps après l’arrêt de la stimulation. De même, lors de la stimulation du ganglion cervical supérieur au cours d’interventions sous anesthésie locale, on a pu constater durant toute la durée de l’excitation une mydriase et une exophtalmie assez prononcées du côté stimulé [55].
Alors que l’on ne retrouve dans la littérature que peu de cas cliniques de syndrome de Pourfour du Petit dont l’origine serait supra-médullaire, les expérimentateurs ont pu obtenir ce syndrome par stimulation chez l’animal des structures supérieures sympathiques, permettant ainsi de situer centres sympathiques et trajet des fibres.
Hensen et Völckers sont les premiers à étudier les effets d’excitation obtenus à partir de la région hypothalamique. Après section transversale de la région thalamique, ils obtiennent un syndrome d’excitation du sympathique cervical (une mydriase avec élargissement de la fente palpébrale) en excitant “le plancher de la surface grise dans la région des tubercules quadrijumeaux et de la commissure postérieure” [24,26].
L’excitation chez le chat d’un point situé à la base du thalamus, derrière la bandelette optique, sur le côté de l’infundibulum (ce centre sympathique correspond à la partie la plus frontale de la région hypothalamique) provoque à la fois une dilatation maximale des pupilles, une ouverture de la fente palpébrale et une rétraction de la paupière interne. L’ablation de l’écorce cérébrale ne modifie pas ces résultats [55,58].
En 1911 Bechterew obtient par stimulation de la paroi interne du thalamus une mydriase, une exophtalmie, une ouverture de la fente palpébrale et une rétraction de la membrane nictitante, résultat non retrouvé après section transversale du thalamus.
Karplus et Kreidl, par excitation électrique du centre qui portera leur nom et qui se situe à la base du thalamus, en arrière de la bandelette optique et en avant de nerf moteur oculaire commun, à la partie externe de l’infundibulum (région du corps de Luys) obtiennent un syndrome bilatéral d’excitation du sympathique oculaire : mydriase, élargissement de la fente palpébrale, rétraction de la membrane nictitante. Ils établissent que l’action de ce centre se fait à la fois par l’intermédiaire de la voie pupillo-dilatatrice et par une inhibition du centre constricteur [26].
En 1927 F.H. Levy confirme les travaux de Karplus et Kreidl et attribue la bilatéralité du syndrome à la diffusion du courant d’excitation au centre controlatéral. En stimulant avec un courant très faible, il observe en effet un syndrome d’excitation sympathique oculaire homolatéral et unilatéral, alors que l’extirpation du ganglion cervical supérieur supprime cette réaction. Il montre qu’en augmentant l’intensité de stimulation la possibilité de réaction à l’excitation cesse en raison d’une lésion du centre, celle-ci étant souvent suivie d’un syndrome de Claude Bernard-Horner qui s’atténue et disparaît les jours suivants. D’autres auteurs (dont Shinosaki Tetsushiro en 1929) confirment ces résultats [26].
A l’instar de Karplus et Kreidl beaucoup de nombreux auteurs observent qu’une dilatation pupillaire maximale peut être provoquée par stimulation électrique de l’hypothalamus [58]. La stimulation d’une région large dans l’hypothalamus ventrolatéral produit une mydriase rapide et maximale due à l’excitation sympathique. Cette réponse est très atténuée par la sympathectomie et n’est pas abolie par la section du nerf optique ni par celle du nerf oculomoteur. Elle existe encore sur une pupille atropinisée [58].
Toutes ces constatations expérimentales sont retrouvées en clinique humaine et servent de base au diagnostic des lésions en cause.